Oddelenie červených rias – rhodophyta. Červené riasy Červené morské riasy 4 písmená

 Živý denník
Živý denník Facebook
Facebook Twitter
TwitterVo veľkých hĺbkach, až 250 metrov, rastú červené riasy, inak nazývaný šarlátový. Rôznofarebné šarlátové rybky v kombinácii s koralmi a žiarivými rybami vytvárajú jedinečnú krásu podmorského sveta. Sú to prevažne veľké riasy, ale napríklad aj mikroskopické červené riasy patria do triedy Banguiaceae.
Prečo môžu červené riasy rásť v takých veľkých hĺbkach? Táto otázka bola položená na Jednotnej štátnej skúške z biológie. Červený pigment umožňuje riasam rásť vo veľkých hĺbkach fykoerytrín. Vďaka nej červené riasy počas fotosyntézy absorbujú zelené, modré, modrofialové lúče spektra. Práve tieto lúče, na rozdiel od červených, sú schopné preniknúť hlboko do vodného stĺpca.
Červené riasy sa vyznačujú asexuálnou reprodukciou spór, ako aj sexuálnou reprodukciou (oogamia); niekedy sa vyskytuje aj vegetatívna reprodukcia časťami talu.
Medzi červené riasy patrí phyllophora, porphyra, gracillaria, ptilote, chondrius a celkovo je to asi päťtisíc druhov.
Fialová- plochá a tenká oválna doska s priemerom do pol metra. Vyznačuje sa iba sexuálnym rozmnožovaním. Mužské reprodukčné bunky nemajú bičíky (spermie). Je to pochopiteľné, pretože vo veľkých hĺbkach pod vodným stĺpcom je ťažké pohybovať sa pomocou bičíka.
Význam rias
Miesto v potravinových reťazcoch, vplyv na prírodu
1. Obrovská masa rias vytvára fytoplanktón a aj v arktických moriach je 20-30 miliónov jedincov na 1 kubický meter vody. Ide o primárne produkty, ktoré tvoria základ potravinového reťazca.
2. Fytoplanktón slúži ako potrava pre zooplanktón (sekundárne produkty), ktoré jedia veľkí morskí obyvatelia, napríklad veľryby. Je zaujímavé, že Thor Heyerdahl počas expedície na plti Kon-Tiki vyskúšal planktónovú „polievku“ a zistil, že je celkom chutná a výživná.
3. Spodné riasy poskytujú úkryt pre ryby a rôzne morské živočíchy a slúžia im aj ako potrava. Kelp napríklad s radosťou žerú morskí ježkovia.
4. Riasy nasýtia oceány a atmosféru kyslíkom.
5. Keď sa však riasy (napríklad Chlamydomonas) počas takzvaného „kvitnutia“ vody masívne rozmnožia, obsah kyslíka v nich klesne a voda sa nasýti toxínmi. Obyvatelia riek zomierajú na nedostatok kyslíka.
Význam pre ľudí
1. Riasy sa v mnohých regiónoch konzumujú ako potrava už tisíce rokov. Obľúbené sú najmä hnedé a červené riasy ako chaluha, undaria, porfýra, hijiki (a sargassum všeobecne).
2. Červené riasy sú zdrojom jódu, najmä niektoré druhy naň bohaté.
3. Červené riasy sú tiež zdrojom agar-agaru, gélotvornej látky, ktorá sa používa v cukrárskom priemysle, na kultiváciu baktérií atď.
4. Pomocou rias sa čistí odpadová voda od fluóru, dusíka a pod., ale aj vzduch od oxidu uhličitého (podarilo sa to Chlamydomonas, Chlorella, Euglena).
5. Riasy sú prídavné látky v potravinách: spirulina, kelp, fucus, ulva, chlorella a iné.
Biologickým významom výskytu súkvetí je zvyšujúca sa pravdepodobnosť opelenia kvetov entomofilných aj anemofilných rastlín. Niet pochýb o tom, že hmyz navštívi oveľa viac kvetov za jednotku času, ak sú zhromaždené v súkvetiach. Okrem toho kvety zhromaždené v kvetenstvách sú nápadnejšie medzi zelenými listami, a nie jednotlivými kvetmi. Mnohé ovisnuté súkvetia sa ľahko kývajú vplyvom pohybu vzduchu, čím sa uľahčuje šírenie peľu.
Súkvetia sú charakteristické pre veľkú väčšinu rastlinných kvetov. Zvyčajne sú kvetenstvo zoskupené v blízkosti vrcholu rastliny na koncoch konárov, ale niekedy, najmä v tropických stromoch, sa objavujú na kmeňoch a hrubých vetvách. Tento jav je známy ako caulifloria. Príkladom je čokoládový strom. Predpokladá sa, že v podmienkach tropického lesa karfiol robí kvety prístupnejšími pre opeľujúci hmyz. Ďalším príkladom karfiolu je strukovina Cercis rohovník, ktorý sa hojne pestuje na Kryme a na Kaukaze.
Súkvetie má hlavnú os alebo os súkvetia a bočné osi, ktoré môžu byť v rôznej miere rozvetvené alebo nerozvetvené. Ich koncové vetvy - pedicely - nesú kvety.
U jednoduché Bočné osi súkvetí nie sú rozvetvené a sú stopkami. Súkvetia, v ktorých sa bočné osi rozvetvujú, sa nazývajú komplexné. Komplexné kvetenstvo môže mať bočné osi prvého, druhého a nasledujúcich rádov.
Hlavná a všetky koncové bočné osy môžu byť zakončené vrcholovými kvetmi, v dôsledku čoho je ich rast obmedzený. Takéto kvetenstvo sa nazýva uzavreté ( sympodiálny), alebo niektoré. V uzavretých súkvetiach sa vrcholové kvety zvyčajne otvárajú skôr ako spodné postranné, a preto sa nazývajú vrcholové kvety.
V otvorených súkvetiach je rast hlavnej osi neobmedzený (t. j. otvorený) a kvety sú umiestnené na strane morfologického hrotu kvitnúceho výhonku. To isté môže platiť aj pre bočné osi. Takéto kvetenstvo sa nazýva otvorené ( monopolný), alebo nedefinované. V otvorených súkvetiach kvety kvitnú postupne zdola nahor, preto sa nazývajú bočné.
2. Monopodial, alebo botryodnye kvetenstvo.
Monopodiálne (otvorené alebo bočné) súkvetia majú hlavnú os, ktorá neustále rastie, a bočné osi nesú kvety. V takomto súkvetí kvitnú najskôr spodné kvety a potom postupne horné. Existujú jednoduché a zložité botryoidné kvetenstvo
Jednoduché botryoidné súkvetia
Štetec - na hlavnej osi alebo osi prvého rádu sú striedavo umiestnené pedicely, ktoré nesú malé kvety (konvalinka, čerešňa vtáčia, náprstník);
Jahňa je hlavnou osou súkvetia, na rozdiel od hrozna visí dole (breza);
Tŕň – na hlavnej predĺženej osi sú sediace kvety (plantain);
Klas - hlavná os súkvetia je veľmi mäsitá, usporiadanie kvetov je ako u klasu (kukurice);
Sh i t o k - kvety sú umiestnené v rovnakej rovine, spodné kvety majú dlhšie stopky ako horné (hruška, jablko, slivka);
Dáždnik - stopky jednotlivých kvetov sú rovnako dlhé a siahajú od hornej časti osi ako lúče dáždnika (cibuľa, čerešňa);
Košík - početné sediace kvety sú umiestnené na skrátenej a tanierovitej zhrubnutej osi. Vonku je obal z listeňových listov (slnečnica, harmanček);
Hlava – blízko seba umiestnené kvety (ďatelina, pŕhľava) sedia na skrátenej a rozšírenej hlavnej osi.
Komplexné botryoidné kvetenstvo
Vyznačujú sa tým, že kvety sú umiestnené na osiach 2., 3. rádu atď.
1. Panicle – na bočných osiach sú jednoduché kefky (orgován, hrozno).
2. Zložitý klások – na hlavnej osi sú umiestnené jednoduché klásky (pšenica, raž, jačmeň).
3. Komplexný dáždnik – na bočných osiach sú jednoduché dáždniky. Jednoduché dáždniky majú zvyčajne svoje zákrovy (súkromné zákrovy) a zákrovy na spodnej časti dáždnika tvoria spoločný zákrov (kôpor, mrkva, aníz).
4. Zložité scutellum (korymbózna metlina) – na bočných osiach sú drobné súkvetia-koryte (jarabina).
V niektorých prípadoch sa pozorujú zmiešané kvetenstvo. Napríklad ovos má metlinové súkvetie, ale namiesto kvetov sú malé klásky, takže sú zmiešané dve súkvetia: metlina a klas.
3. Sympodiálne alebo cymoidné súkvetia. Morfológia ovocia.
Sympodiálne (cymoid) sú tie súkvetia, v ktorých sa stopka (hlavná os) končí kvetom a v jeho raste pokračujú horné postranné výhonky alebo postranné výhonky, ktoré zase pokračujú v raste rovnakým spôsobom. Cymoidné kvetenstvo zahŕňa nasledujúce:
1. Vidlička, alebo dicházia. Hlavná os končí kvetom, rast stopky pokračuje s protiľahlými postrannými púčikmi nižšie, tvoriac kvety atď.
2. Pleiocházium alebo falošný dáždnik - rast stopky končí kvetom a pokračuje zvinutým pod postrannými púčikmi, ktoré tvoria kvety (euphorbia).
3. Kučera - rast stopky končí kvetom a pokračuje bočným púčikom, ktorý tvorí kvet a tak ďalej opakovane, pričom všetky kvety smerujú jedným smerom (zemiak, nezábudka, skorocel).
Morfológia ovocia.
Po oplodnení sa steny piestikového vaječníka vyvinú na oplodie (perikarp). Oplodie je stena plodu obklopujúca semeno, ktorá sa tvorí súčasne s oplodím. Oplodie sa zvyčajne skladá z troch vrstiev:
1) externé ( exokarp), niekedy pokryté rôznymi výrastkami (perutýn javorový, prívesy lopúcha);
2) stred ( mezokarp), ktorý tvorí dužinu šťavnatého ovocia, obsahujúcu veľa cukru (slivka, čerešňa) alebo oleja (olivový);
3) interné ( endokarp),často sa mení na vrstvu šťavnatého ovocia
kamenné bunky (kôstka slivky, kôstka čerešne) alebo do šťavnatej dužiny (citrón).
Všetky tri zóny sú jasne rozlíšiteľné. Napríklad v plodoch čerešne je tenká kožovitá vonkajšia vrstva - exokarp, jedlá šťavnatá dužina ovocia je mezokarp, tvrdá kôstka kamenistého tkaniva obklopujúca jediné semeno - endokarp.
Nezrelé plody majú zvyčajne zelenú farbu, bunky exokarp a mezokarp obsahujú chlorofyl a fotosyntetizujú. Dozrievaním plody strácajú zelenú farbu a často získavajú jasnú farbu, ktorú spôsobujú buď antokyány v bunkovej šťave (čerešne, hrozno, čučoriedky) alebo chromoplasty (jarabina, paradajka).
V niektorých rastlinných druhoch sa ovocie tvorí nielen z vaječníka piestika, ale aj z nádobky alebo periantu. Takéto ovocie sa nazýva falošné. Napríklad bobuľovitý falošný plod jahody je prerastená šťavnatá farebná nádoba, na ktorej sú malé suché skutočné plody (nažky), vytvorené z vaječníkov piestikov. Zo zarastenej nádoby sa tvoria plody šípok, jabloní a hrušiek.
Pri niektorých rastlinných druhoch sa plod tvorí z niekoľkých piestikov. Takéto ovocie sa nazýva prefabrikované, alebo komplexné (zložené ovocie malín, černíc, masliakov).
4. Klasifikácia ovocia. Všetky druhy ovocia sú rozdelené do nasledujúcich typov.
Nevýrazné suché ovocie
1. Orech alebo orech má drevnaté oplodie obsahujúce jedno voľné semeno. Napríklad v lieske, dube, buku, konope.
2. Nažka má kožovité oplodie, ktoré obsahuje jedno voľne ležiace semeno. Napríklad v slnečnici, púpave, rumančeku a ďalších druhoch z čeľade astrovité.
3. Obilka má kožovité oplodie, ktoré obsahuje jedno semienko zrastené s oplodím. Napríklad v pšenici, jačmeni a iných druhoch z čeľade obilnín.
4. Perutýn má oplodie v tvare orecha alebo nažky, ktoré prerástlo do jedného alebo viacerých krídlových príveskov. Napríklad javor, breza, brest, jaseň.
5. U niektorých druhov má suchý, nevýrazný plod muchu - chumáč chlpov. Napríklad púpava.
Otváranie sušeného ovocia
1. Leták - škatuľkovitý, jednomiestny, viacsemenný plod, tvorený jedným plodolistom, otvárajúcim sa pozdĺž ventrálneho stehu od vrcholu k základni.
Napríklad pri bavlníkoch, druhoch čeľade pryskyřníkovitých a ružovitých. Pivonka vytvára veľa lístkov z piestikov jedného kvetu.
2. Bob – škatuľkovitý, jednomiestny, často viacsemenný plod, tvorený jedným plodolistom, otvárajúcim sa pozdĺž ventrálnych a chrbtových stehov od vrcholu k základni. Napríklad v hrachu a iných druhoch z čeľade strukovinových. U niektorých druhov sa fazuľové chlopne pri otvorení špirálovito skrúcajú a rozhadzujú semená (napríklad v žltej akácii). Pri niektorých druhoch sú fazule jednosemenné (napríklad v sainfone) a potom sa neotvárajú.
3. Lusk a silique sú škatuľkovité, dvojjazérne, viacsemenné plody tvorené dvomi plodolistami. V strede plodu zhora nadol je membránová prepážka, ku ktorej okrajom sú pripevnené semená. Ovocie sa otvára dvoma chlopňami pozdĺž dvoch švíkov od základne po vrch. Ventily odpadnú, ale prepážka so semenami zostane. Struk je dlhý a úzky plod - dĺžka je 4-krát alebo viackrát väčšia ako šírka (napríklad horčica) a struk je krátky a široký (napríklad pastierska kabelka).
4. Tobolka - mnoholodný (zriedka unilokulárny) mnohosemenný plod tvorený niekoľkými plodolistami. Škatule sa otvárajú rôzne, buď chlopňami (napríklad bavlna, durman, ľan, čaj, ľalia, ricínové bôby), alebo zúbkami na vrchu škatuľky (napríklad pri mnohých druhoch čeľade klinčekovité), alebo s otvormi (napr. pri niektorých druhoch maku a zvončeka), buď s vrchnákom na vrchu škatuľky (napr. v kurníku), alebo s bočnými štrbinami (napr. v okra).
U niektorých druhov sa suché viacsemenné plody (struky, fazuľa) po dozretí priečne rozpadávajú na jednosemenné segmenty (napríklad pri divokej reďkovke). Takéto plody sa nazývajú kĺbové.
U iných druhov sa suché viacsemenné plody po dozretí pozdĺžne rozpadajú na jednotlivé jednosemenné segmenty, napríklad pri javoroch a iných druhoch z čeľade javorovitých, ako aj pri čeľadiach Apiaceae, Lamiaceae, Malvaceae a niektorých ďalších. Takéto ovocie sa nazýva zlomkové.
Šťavnaté ovocie
1. Bobuľa je šťavnatý, zvyčajne farebný, viacsemenný plod tvorený jedným alebo viacerými plodolistami. Semená v bobule sú vložené do šťavnatej dužiny. Napríklad hrozno, ríbezle, egreše, belladonna, čučoriedky, brusnice, zemiaky, paradajka, uhorka, melón, melón, tekvica, citrón, mandarínka, pomaranč (uhorka, tekvica, vodný melón, melón sú falošné bobule, pretože vonkajšia časť dužinaté ovocie, ktoré vytvorili z nádoby).
2. Kôstkovica - zvyčajne šťavnatý a farebný, jednosemenný alebo viacsemenný plod, tvorený jedným alebo viacerými plodolistami. Endokarp kôstkovice má vzhľad kameňa. Napríklad slivky, čerešne, marhule, drieň, olivy. Viackôstkové kôstkovice - v baze čiernej, rakytník preháňavý. Plody vlašského orecha sú kôstkovice, ale jeho mezokarp nie je šťavnatý; samotný takzvaný orech je semenom plodu. Mezokarp mandľovej kôstkovice tiež nie je šťavnatý, zatiaľ čo kôstkovina kokosovej palmy je vláknitá.
Plodnosť krytosemenných rastlín je veľmi vysoká. Jedna rastlina bielej quinoy teda produkuje asi 100 000 semien ročne, kurník - ≈ 500 000, quinoa - ≈ 750 000,
topole ≈ 27 000 000 semien.
5. Neplodnosť. Distribúcia ovocia a semien.
U niektorých druhov sa plod tvorí súkvetie. Táto formácia sa nazýva neplodnosť. Napríklad plodom repy je guľa, plodom moruše je „bobule“, plodom viniča je „bobule“. Vňať moruše sú zrastené nepravé plody tvorené previsnutými periantami.
V niektorých odrodách kultúrnych rastlín sa plody vyvíjajú bez opelenia a oplodnenia, a teda bez semien. Tento jav tvorby plodov bez semien sa nazýva partenokarpia. Partenokarpia sa pozoruje u niektorých odrôd jabloní, hrušiek („bezsemenných“), hrozna (hrozienka – sušené hrozno bez jadierok), egrešov, mandarínok, pomarančov, citrónov, fíg, japonského tomelu atď. U niektorých odrôd môže byť partenokarpia spôsobená dráždenie blizny cudzím peľom (opelenie hrušky peľom jabĺk, paradajky peľom zemiakov, baklažánu peľom paradajok), niektorými chemikáliami, bodnutím hmyzom, pálením horúcim drôtom a pod. Plody bez semien sú zvyčajne menšie ako plody so semenami a , preto produkujú menšie výnosy. V praxi sú však plody bez semien spotrebiteľmi veľmi cenené. Bezsemenné odrody sa môžu množiť iba vegetatívne.
Distribúcia ovocia a semien
Semená a plody krytosemenných rastlín vykazujú mnoho rôznych adaptácií na šírenie. Väčšina rastlinných druhov sa prispôsobila roznášaniu semien a plodov vetrom (anemochory). Semená bavlny, osiky, topoľa, vŕby, púpavy atď. majú zvláštne dlhé chĺpky a vločky, ktoré uľahčujú ich rozptýlenie vetrom, často na veľké vzdialenosti. Plody javora, brezy, brestu, jaseňa, jelše, semená borovice, smreka a pod. K rozptylu vetra prispieva aj krycí list súkvetia lipy. Semená mnohých druhov z čeľadí vresovcov, horcov, orchideí atď., sú také malé a ľahké, že ich vietor ako prach unáša na veľmi veľké vzdialenosti. Rastliny ťavej trávy, kurai, chrobáka atď., ktoré majú guľovitý tvar, sú po dozretí plodov odlomené vetrom z podzemnej časti a váľajú sa po stepi na veľké vzdialenosti, pričom často vytvárajú celé pohyblivé šachty, rozptyľujú sa ich semená po ceste. Tieto rastliny sú bežne známe ako tumbleweeds.
Semená a plody vodných rastlín sú rozptýlené vodou (hydrochória). Dažďová voda (najmä po daždi), voda z potokov a riek zohráva významnú úlohu pri prenose semien a plodov rastlín pestovaných na súši.
Živočíchy hrajú hlavnú úlohu pri šírení semien. Šťavnaté plody jedia vtáky (ornithochory) a v menšej miere aj iné živočíchy (zoochory). Ich semená, ktoré prechádzajú tráviacim traktom zvieraťa, nie sú poškodené a nestrácajú svoju životaschopnosť. Spolu s exkrementmi zvierat semená padajú na zem a vyklíčia.
Semená teda často klíčia v značnej vzdialenosti od materskej rastliny. V niektorých druhoch rastlín (lopúch, mrkva, lopúch, suchý zips) majú semená a plody rôzne pripevnenia alebo priľnavé povrchy a priľnú k srsti okoloidúceho zvieraťa alebo sa prilepia na perie vtákov a sú tak transportované. Malé semená šíria mravce a môžu sa tiež prilepiť na kopytá zvierat v špine a byť takto prenášané.
Niektoré druhy rastlín si vyvinuli schopnosť samostatne rozptýliť svoje semená (autochória). V žltej akácii sa teda ventily dozretých bôbov otvárajú a špirálovito otáčajú takou silou a rýchlosťou, že semená prítomné vo fazuli sa silou rozptyľujú na všetky strany. Toto otváranie plodov v žltej akácii sa vysvetľuje tým, že keď dozrieva fazuľa, vonkajšia a vnútorná vrstva chlopní sa nerovnomerne sťahujú a medzi nimi vzniká a rastie napätie, ktoré spôsobuje krútenie a trasenie chlopní pri otváraní zrnka.
V „šialenej“ uhorke, ktorej zrelé ovocie sa odlomí od stopky a semená sa vyhodia do výsledného otvoru spolu s tekutým obsahom. To sa deje s takou silou, že semená zvyčajne padajú niekoľko metrov ďaleko. Toto vyvrhnutie tekutiny a semien sa vysvetľuje skutočnosťou, že ako ovocie dozrieva, tlak sa zvyšuje a v čase dozrievania ovocia dosahuje vysoké napätie.
TÉMA LEKCIE 27 : Ranunculaceae (Ranunculaceae). maková rodina (Papaveracea)
1. Čeľaď Ranunculaceae. 2. Maková rodinka. S. 379 – 383
Rad Ranunculaceae.Čeľaď Ranunculaceae (2.000)
Asi 2000 druhov. Druhy čeľade Ranunculaceae sú bežné v chladných miernych a subtropických (horských) oblastiach. V regióne Smolensk rastie 32 druhov. V trópoch sa rastliny tejto čeľade vyskytujú výnimočne. Formy života sú predovšetkým viacročné bylinné rastliny, kry, trpasličí kríky a vinič sú zriedkavé.
Listy sú jednoduché, bez paličiek; Tvar listovej dosky je rôzny. Listy môžu byť buď celé (jarná tráva), alebo rozrezané v rôznej miere (sasanka lesná, pečeňovník ušľachtilý, kolumbína). Usporiadanie listov je pravidelné.
Mnohé masliaky majú upravené podzemné výhonky (podzemok veternice a konské šišky jarnej číry).
Kvety môžu byť jednotlivé (rod sasanka) alebo zhromaždené v súkvetiach (rod ranunculus, kolumbína, nevädza). Ranunculaceae sú mimoriadne rozmanité v štruktúre kvetov.
Rozmanitosť v štruktúre kvetov môže byť vyjadrená sledovaním niekoľkých evolučných sérií.
1. Kvety s neurčitým počtom členov (u sasanky obyčajná môže mať 5-8 listov) → kvety s určitým, jasne stanoveným počtom členov (bojovník rodu).
2. Okvietnica jednoduchá (rod sasanka, rod columbine) → okvetie dvojité (rod Ranunculus)
3. plodolistov z veľkého počtu na jeden
4. gynoecium z apokarpného na synkarpné
5. kvety od aktinomorfných kvetov po zygomorfné (rod zápasník, rod akonita
6. os kvetu je predĺžená, takže kvetinové členy sú usporiadané do špirály → os kvetu je takmer plochá, takže kvetinové členy sú usporiadané do kruhu.
7. kvety sú nešpecializované, entomofilné (často opeľujú muchy) → kvety sú špecializované, entomofilné (rod zápasník, rod Larkspur).
Mnohé druhy masliakov sa vyznačujú tým staminódy- značne zmenšené a upravené okvetné lístky (alebo tyčinky), ktoré sa menia na nektáriá. Iba rastlina bazalka nemá nektary.
Väčšina druhov má piestikov veľa, sú voľné a umiestnené na predĺženej osi (rody masliaka, pečeňovník, nechtík). Menej často je počet piestikov obmedzený: 3-1. Vaječník je vždy unilokulárny a nadradený. Obvykle je veľa vajíčok, ale sú druhy, ktoré majú v piestiku 1 vajíčko. Plody sú letáky alebo orechy.
Môže existovať rôzny počet tyčiniek. Najčastejšie sú kvety s veľkým a neurčitým počtom tyčiniek, u niektorých druhov môže byť od 3 do 1 tyčiniek.
nechtík močiarny – * Р 5 А ¥ G ¥
V závislosti od druhu ovocia sa v čeľade Ranunculaceae rozlišujú dve podčeľade:
· zimomriavky (rody plavky, zápasník, škovránok), majúce letákové ovocie
· Ranunculaceae (rody Ranunculus, Anemone, Basil), majúce orieškové plody
Alkaloidy sú veľmi bežné vo všetkých rastlinách tejto čeľade. Ranunculaceae sú jedovaté, nie sú to živné rastliny, ale pre množstvo alkaloidov sa rastliny využívajú ako liečivá. V morfologicky nešpecializovaných druhoch (rod bazalka, nechtík a lipkavec) sa syntetizujú jednoduché alkaloidy, pri pokročilých a špecializovaných rodoch komplexné alkaloidy (rastliny z rodu akonit a larkspur).
Z rôznych druhov masliakov sa v kardiologickej praxi používa 20 alkaloidov. Alkaloidy všetkých druhov masliakov sú jedovaté, najjedovatejšie patria do čeľade akonitových.
Adonis jarný, retikulát retikulát a bórax (alebo akonit) sú dôležité liečivé rastliny.
Alkaloidy v jarnom adonise vyvolávajú účinok podobný kurare a používajú sa na uvoľnenie svalov.
Alkaloidy larkspur reticularis sa používajú v nervovej klinike pri ochoreniach spojených s poruchami motorických funkcií
Pri radikulitíde sa odporúča tinktúra z byliny Djungarian akonit, ktorá je súčasťou lieku "Akofit".
Vzácny a chránený druh: plavec európsky, vrana ostnatá, zápasník severský.
Objednajte si Poppy.Poppy family (250)
Rastliny tejto čeľade sú rozšírené hlavne v tropických a subtropických oblastiach. Zriedkavo sa vyskytuje v miernom pásme. Na území regiónu Smolensk sa vyskytujú 2 druhy - skorocel väčší a mak.
Mak - bylinné trvalky, niekedy ročné rastliny. Prítomnosť letničiek sa považuje za evolučne mladú vlastnosť. Usporiadanie listov je striedavé, listy sú jednoduché, bez paličiek, môžu byť celé alebo rozrezané.
Kvety maku sú často veľké a aktinomorfné:
* Ca 2 Co 2 + 2 A ¥ G ( ¥ )
Sepaly spravidla opadávajú, keď sa kvet otvorí. Gynoecium pozostáva z niekoľkých plodolistov, ktoré spolu zrastú a vytvárajú parakarpné gynoecium. Plodom je suchá tobolka obalená klinčekmi. Existuje veľa vajíčok, plody sú malé a obsahujú tuky ako rezervný materiál.
Nápadným anatomickým a biochemickým znakom maku je prítomnosť laticiferov, v ktorých sa syntetizuje mliečna šťava komplexnej štruktúry. Obsahuje rôzne izochinolínové alkaloidy. Mliečna šťava a alkaloidy maku sú široko používané v medicíne.
Osobitný význam nadobudol rod mak, ktorý je hlavnou surovinou na výrobu ópia. Ópium obsahuje viac ako 20 alkaloidov vrátane morfínu, kodeínu, narkotínu a papaverínu. Niektoré alkaloidy majú silný anestetický účinok, ale sú návykové. Papaverín sa používa ako spazmolytikum pri angíne pectoris a bronchiálnej astme. U nás sa mak uspávajúci nepestuje. Určité množstvo omamných alkaloidov obsahujú aj olejnaté a okrasné odrody maku.
Skvelý celandín
Kvety sú malé, zhromaždené v dáždnikovom kvetenstve: * Ca 2 Co 2 +2 A ¥ G (¥)
Jeho pomarančová mliečna šťava spolu s alkaloidmi obsahuje flavonoidy, triesloviny, saponíny, organické kyseliny a vitamíny. Bylina skorocelu sa používa ako choleretikum a antibakteriálne činidlo pri ochoreniach pečene a žlčníka. Skorocel väčší je jedovatá rastlina.
Liečivo „Sangviritrin“ sa vyrába z skorocelovej byliny na vyplachovanie úst a hrdla pri zápale, ako aj na umývanie hnisavých rán.
Mak žltý sa používa na liečebné účely a získava sa nie narkotické antitusikum glaucín hydrochlorid. Na liečebné účely sa využíva aj makaka maloplodá a makaka srdcovitý.
LEKCIA 28: Čeľaď Rosaceae.
1. Čeľaď Rosaceae, všeobecná charakteristika.
2. Podčeľaď Spiraea. Podrodina ružová.
3. Podrodina jabĺk. Podrodina sliviek.
1. Čeľaď Rosaceae (3500)
Zástupcovia tejto rodiny sú distribuovaní mimo tropických oblastí severnej pologule a nachádzajú sa v rôznych fytocenózach. V regióne Smolensk rastie 56 druhov Rosaceae. Forma života: stromy, kríky, byliny (jednoročné a viacročné).
Listy sú jednoduché (celé alebo členité), ako aj zložité (párové a nepárovité, trojpočetné), veľmi často sa vyskytujú palisty. Usporiadanie listov je striedavé, zriedka opačné. Niektoré druhy majú tŕne (metamorfózy epidermis).
Biochemické vlastnosti Rosaceae:
· nedochádza k nešpecializovanému sekundárnemu metabolizmu. To sa prejavuje pri úplnej absencii alkaloidov a toxínov. Esenciálne oleje vznikajú zriedkavo a vždy majú jednoduché zloženie a štruktúru (s výnimkou rodu Rose)
Štruktúra kvetov Rosaceae nie je vysoko špecializovaná na opeľovanie hmyzom. Kvety sú jednotlivé alebo zhromaždené v súkvetiach rôznych typov. Kvety sú aktinomorfné, okvetie je zložité - zvyčajne je päť sepalov a okvetných lístkov. Pomerne často má kvet dvojnásobný počet sepalov, v takom prípade sa vytvorí prvý kruh sepalov menejcenný. Existuje veľa tyčiniek, 2-4 krát viac ako okvetné lístky. Počet plodolistov je buď neurčitý, alebo prísne stanovený.
Charakteristickým znakom kvetu je prítomnosť hypanthium- zvláštny útvar vytvorený zo zarastenej nádobky a s ňou zrastených základov sepalov, okvetných lístkov a tyčiniek. Tvar hypantia môže byť konvexný, tanierovitý alebo konkávny. Hypanthium chráni gynoecium pred environmentálnymi faktormi. Zároveň hrá dôležitú úlohu pri tvorbe plodov. Môže rásť a podieľať sa na tvorbe mezokarpu a endokarpu. Šťavnatá dužina šípok, jabloní, hrušiek a sliviek je teda prerastené hypanthium. Okrem toho hypanthium zabezpečuje komunikáciu so zvieratami, ktoré roznášajú ovocie (exo- a endochória).
Plody Rosaceae majú veľmi rôznorodú štruktúru, čo poskytuje rôzne spôsoby distribúcie. Zlepšenie metód distribúcie je jedným z hlavných smerov evolúcie v rámci rodiny.
Rosaceae veľmi často produkujú falošné plody (jablko v jabloni a jablko v horskom jaseni) a komplexné ovocie (polyorech v jahodách a kôstkovice v malinách).
Čeľaď Rosaceae zahŕňa štyri podčeľade: spireaceae, roseaceae, jablko a slivku.
2. Podčeľaď Spiraea. Podrodina ružová.
Podčeľaď Spiraea (180)
Ide o najprimitívnejšiu podčeľaď, ktorú predstavujú kríky, menej často stromy a veľmi zriedkavo trvalé rizomatózne byliny. Kvety sú malé, zhromažďujú sa buď v metlinách, alebo v hroznovitých súkvetiach alebo v corymboch. Okvetné lístky koruny sú biele, menej často svetloružové. Zástupcovia tejto čeľade sa vyznačujú konkávnou alebo takmer plochou nádobou (hypanthium) a plody sú suché - viaclisté (kvintalisté). Gynoecium je apokarpné, zvyčajne pozostáva z 2-5 plodolistov.
Do čeľade patria rody Spirea, Meadowsweet a Jarabina.
Ústredným rodom je rod Spiraea, rozšírený na Sibíri a na Ďalekom východe. Spiraea sú kry s jednoduchými listami, bez palistov. Kvety sú zvyčajne biele alebo ružové, v metlinovitých, corymbose alebo dáždnikových súkvetiach. Plody sú viaclisté.
Rod lúčna, druh lúčna. Je to veľká trváca rastlina vysoká až 2 m, s krátkym podzemkom. Listy sú prerušovane sperené, zhora tmavozelené, zospodu tenké bielo plstnaté dospievanie. Pri trení listy vydávajú štipľavý zápach. Kvety sú žltkastobiele a zhromaždené v hustých panikulárnych kvetenstvách. Plody sú jednosemenné lístočky. V ľudovom liečiteľstve sa odpradávna používala lúčna medovka ako adstringens.
* Ca (5) Co 5 A ¥ G 6-10
Podčeľaď Rosaceae (1 700)
Bylinné rastliny sú rozšírené v rôznych fytocenózach od tundry až po horské trópy. Kvety sa zhromažďujú v corymbose alebo racemose dichasia, menej často osamelé kvety.
psia ruža – * Ca (5) Co 5 A ¥ G ¥
Gynoecium je apokarpné, polymérne, ale vaječník obsahuje vždy jedno, zriedkavo dve vajíčka.
Plody sú rozmanité, ale najčastejšie sú to kôstkovice a mnohoraké. Mnohé druhy tejto čeľade majú šťavnaté komplexné plody, na tvorbe ktorých sa podieľa hypanthium.
Mnohé rastliny tejto čeľade sa vyznačujú rozmnožovaním pomocou odnoží, stolonov alebo úponkov.
Ako liečivá sa používajú látky sekundárneho metabolizmu ruží.
1. malina obyčajná - jej plody obsahujú značné množstvo kyseliny salicylovej, ktorá podmieňuje ich diaforetický a antipyretický účinok proti prechladnutiu.
2. palina - kvet nemá korunu, kalich je fialový a 4-dielny. Odvar z podzemku sa používa ako hemostatický prostriedok na krvácanie z maternice a hemoroidov.
3. Potentilla erecta (Uzik, Kalgan). Vyznačuje sa 4-členným okvetím a kalichom s podkalichom. Galangalová tinktúra (25g podzemku na 500ml vodky) sa používa pri črevných kataroch a úplavici.
4. skorocel močiarny (ľudový názov - dekop) - odvar z podzemku sa používa pri ochoreniach kĺbov a žltačke. Používa sa tiež ako adstringens, hemostatikum a diaforetikum.
5. Škorica šípková – obsahuje veľké množstvo vitamínu C, ďalej vitamíny B2, K, karotén a kyselinu citrónovú. Kondenzovaný vodný extrakt zo šípky s cukrom (holosas) sa používa ako choleretikum.
3. Podrodina jabĺk. Podrodina sliviek.
ODDELENIE ČERVENÝCH RIAS tvoria takmer výlučne morské riasy. životný cyklus - úplná absencia bičíkových štádií; špeciálna forma oogamného sexuálneho procesu. 2
 Pigmenty chlorofyl „a“ a „b“, karotenoidy (karotén, zeaxantín, anteraxantín, kryptoxantín, luteín, neoxantín) fykobilíny: fykoerytríny (červené) fykokyaníny a alofykokyanín (modrá) farba oceľovo-modrého talu od modrej po oceľovomodrú nadbytok fykokyanínu) 3
Pigmenty chlorofyl „a“ a „b“, karotenoidy (karotén, zeaxantín, anteraxantín, kryptoxantín, luteín, neoxantín) fykobilíny: fykoerytríny (červené) fykokyaníny a alofykokyanín (modrá) farba oceľovo-modrého talu od modrej po oceľovomodrú nadbytok fykokyanínu) 3
 Chloroplasty sú obalom dvoch membrán, jednotlivých tylakoidov, na povrchu fykobilizómu. Rezervným produktom je polysacharid „purpurový škrob“, ktorý získava od jódu hnedočervenú farbu. Fialové škrobové zrná sú uložené v cytoplazme vždy bez spojenia s pyrenoidmi a chromatofórmi. 4
Chloroplasty sú obalom dvoch membrán, jednotlivých tylakoidov, na povrchu fykobilizómu. Rezervným produktom je polysacharid „purpurový škrob“, ktorý získava od jódu hnedočervenú farbu. Fialové škrobové zrná sú uložené v cytoplazme vždy bez spojenia s pyrenoidmi a chromatofórmi. 4
 Štruktúra talu je jednobunková kokoidná forma (porphyridium), heterotrichná a vo forme rozvetvených filamentov pripevnených k substrátu pomocou rizoidov. pseudoparenchymatózne stielky, prepletanie bočných vetiev Lamelárne stielky parenchymatóznej štruktúry (porfýr). 5
Štruktúra talu je jednobunková kokoidná forma (porphyridium), heterotrichná a vo forme rozvetvených filamentov pripevnených k substrátu pomocou rizoidov. pseudoparenchymatózne stielky, prepletanie bočných vetiev Lamelárne stielky parenchymatóznej štruktúry (porfýr). 5
 Bunka je pokrytá pektínovým obalom - hemicelulózové zložky napučiavajú a v stenách sa ukladá vápno. Bunky sú jedno- a viacjadrové.Chromatofory sú parietálne, početné, vo forme zŕn alebo doštičiek. 6
Bunka je pokrytá pektínovým obalom - hemicelulózové zložky napučiavajú a v stenách sa ukladá vápno. Bunky sú jedno- a viacjadrové.Chromatofory sú parietálne, početné, vo forme zŕn alebo doštičiek. 6
 Nepohlavné rozmnožovanie cez nepohyblivé bunky monospóry - jedna vo výtrusnici Tetraspory - štyri tetraspóry - na diploidných nepohlavných rastlinách - sporofytoch. V sporangiách sa meióza vyskytuje pred tvorbou tetraspór. 7
Nepohlavné rozmnožovanie cez nepohyblivé bunky monospóry - jedna vo výtrusnici Tetraspory - štyri tetraspóry - na diploidných nepohlavných rastlinách - sporofytoch. V sporangiách sa meióza vyskytuje pred tvorbou tetraspór. 7
 Pohlavný výbežok Oogamný Ženský orgán - vo väčšine prípadov karpogón - z rozšírenej bazálnej časti - brucho - vajíčko a výbežok - trichogyna. Carpogon sa vyvíja na špeciálnej krátkej karpogonálnej vetve. 8
Pohlavný výbežok Oogamný Ženský orgán - vo väčšine prípadov karpogón - z rozšírenej bazálnej časti - brucho - vajíčko a výbežok - trichogyna. Carpogon sa vyvíja na špeciálnej krátkej karpogonálnej vetve. 8
 Antherídia - malé bezfarebné bunky obsahujúce bičíkovité spermie, spermie sú pasívne transportované vodnými prúdmi a priľnú k trichogýnu. V mieste kontaktu sa spermie a trichogýny rozpúšťajú ich steny, jadro spermie sa pohybuje po trichogýne v brušnej časti karpogónu, spája sa s 9.
Antherídia - malé bezfarebné bunky obsahujúce bičíkovité spermie, spermie sú pasívne transportované vodnými prúdmi a priľnú k trichogýnu. V mieste kontaktu sa spermie a trichogýny rozpúšťajú ich steny, jadro spermie sa pohybuje po trichogýne v brušnej časti karpogónu, spája sa s 9.
 tvorba karpogónov Po oplodnení je bazálna časť karpogónu oddelená prepážkou od trichogýny - odumiera Vývoj - tvorba karpospór obsah zygoty (oplodneného karpogónu) 1. priamo sa delí za vzniku nepohyblivých holých spór - karpospór, 10
tvorba karpogónov Po oplodnení je bazálna časť karpogónu oddelená prepážkou od trichogýny - odumiera Vývoj - tvorba karpospór obsah zygoty (oplodneného karpogónu) 1. priamo sa delí za vzniku nepohyblivých holých spór - karpospór, 10
 tvorba karpospór 2. z oplodneného karpogónu vyrastajú vetviace nitky - gonimoblasty, ich bunky sa menia na karposporangiá, produkujúce po jednom karpospore. jedenásť
tvorba karpospór 2. z oplodneného karpogónu vyrastajú vetviace nitky - gonimoblasty, ich bunky sa menia na karposporangiá, produkujúce po jednom karpospore. jedenásť
 tvorba karpospór vo väčšine gonimoblastov sa nevyvíja priamo z bruška oplodneného karpogónu z acoxylárnych buniek. môžu byť odstránené z karpogonu alebo sú umiestnené na stélku v tesnej blízkosti, keď sú pomocné bunky odstránené z karpogonu, spojivové bunky alebo oblasti rastú z jeho brucha po oplodnení y e, závity. 12
tvorba karpospór vo väčšine gonimoblastov sa nevyvíja priamo z bruška oplodneného karpogónu z acoxylárnych buniek. môžu byť odstránené z karpogonu alebo sú umiestnené na stélku v tesnej blízkosti, keď sú pomocné bunky odstránené z karpogonu, spojivové bunky alebo oblasti rastú z jeho brucha po oplodnení y e, závity. 12
 bunky oblasticových filamentov obsahujú diploidné jadrá. Oblastové vlákna rastú smerom k pomocným bunkám, v mieste kontaktu sa membrány rozpustia a vytvorí sa komunikácia medzi bunkou oblasticového vlákna a pomocnou bunkou. Táto fúzia buniek nie je sprevádzaná fúziou ich jadier (diploidné bunkové jadro ooblastického vlákna a haploidné jadro pomocného vlákna). fúzia s pomocnou bunkou stimuluje delenie diploidného jadra oblastem filamentnej bunky a vývoj gonimoblastov obsahuje diploidné jadrá a produkujú diploidné karpospory Gonimoblasty - špeciálna generácia - 13
bunky oblasticových filamentov obsahujú diploidné jadrá. Oblastové vlákna rastú smerom k pomocným bunkám, v mieste kontaktu sa membrány rozpustia a vytvorí sa komunikácia medzi bunkou oblasticového vlákna a pomocnou bunkou. Táto fúzia buniek nie je sprevádzaná fúziou ich jadier (diploidné bunkové jadro ooblastického vlákna a haploidné jadro pomocného vlákna). fúzia s pomocnou bunkou stimuluje delenie diploidného jadra oblastem filamentnej bunky a vývoj gonimoblastov obsahuje diploidné jadrá a produkujú diploidné karpospory Gonimoblasty - špeciálna generácia - 13
 V najviac organizovaných červených riasach sa pomocné bunky vyvíjajú až po oplodnení karpogona a v jeho bezprostrednej blízkosti. Kombinácia pomocnej bunky (alebo buniek) s karpogónom sa špeciálne nazýva prokarp. Nie je potrebný vznik dlhých ooblastických filamentov spájajúcich karpogón a pomocné bunky, pomocná bunka jednoducho splynie s bruškom oplodneného karpogónu, po čom sa z nej vyvinú gonimoblasty s karpospórami.Karposporangiá sa často nachádzajú v úzkych skupinách - cystokarpy, pokrytá pseudoparenchymatóznou membránou vyvíjajúcou sa z buniek susediacich s karpogónom. 14
V najviac organizovaných červených riasach sa pomocné bunky vyvíjajú až po oplodnení karpogona a v jeho bezprostrednej blízkosti. Kombinácia pomocnej bunky (alebo buniek) s karpogónom sa špeciálne nazýva prokarp. Nie je potrebný vznik dlhých ooblastických filamentov spájajúcich karpogón a pomocné bunky, pomocná bunka jednoducho splynie s bruškom oplodneného karpogónu, po čom sa z nej vyvinú gonimoblasty s karpospórami.Karposporangiá sa často nachádzajú v úzkych skupinách - cystokarpy, pokrytá pseudoparenchymatóznou membránou vyvíjajúcou sa z buniek susediacich s karpogónom. 14
 15
15

 CLASS BANGUY Bunky často s hviezdicovým chromatoforom a pyrenoidom. Póry medzi bunkami zvyčajne chýbajú. Karpogón bez trichogýnu, po oplodnení sa obsah karpogónu priamo rozdelí na karpospóry. Nepohlavné rozmnožovanie – monospóry 17
CLASS BANGUY Bunky často s hviezdicovým chromatoforom a pyrenoidom. Póry medzi bunkami zvyčajne chýbajú. Karpogón bez trichogýnu, po oplodnení sa obsah karpogónu priamo rozdelí na karpospóry. Nepohlavné rozmnožovanie – monospóry 17

 19
19
 rod compsopogon Sladkovodný rozšírený v trópoch zavlečený s akvarijnými rastlinami Thallus heterotrichous Rozmnožovanie monospórami. Monosporangia sú oddelené šikmou prepážkou od ktorejkoľvek bunky talu a majú zrnitý obsah z množstva purpurového škrobu, ktorý sa mení na jednu nehybnú nahú monospóru 20
rod compsopogon Sladkovodný rozšírený v trópoch zavlečený s akvarijnými rastlinami Thallus heterotrichous Rozmnožovanie monospórami. Monosporangia sú oddelené šikmou prepážkou od ktorejkoľvek bunky talu a majú zrnitý obsah z množstva purpurového škrobu, ktorý sa mení na jednu nehybnú nahú monospóru 20
 21
21

 TRIEDA FLORIDA Bunky väčšinou s parietálnymi chromatofórmi bez pyrenoidov.Medzi bunkami sú póry. Carpogon s trichogynom. Po oplodnení sa gonimoblasty vyvíjajú buď priamo z bruška oplodneného karpogónu, alebo z pomocných buniek po ich splynutí s oblasťovými vláknami. Nepohlavné rozmnožovanie je väčšinou pomocou tetraspór. Gonimoblasty nesúce karposporangiá sa považujú za špeciálnu generáciu - karposporofyt. Podrobnosti o vývoji karposporofytu (z oplodneného karpogónu alebo z pomocných buniek), ako aj čas diferenciácie pomocných buniek (pred alebo po oplodnení), ich poloha na stéli a pod. floridia do šiestich rádov 23
TRIEDA FLORIDA Bunky väčšinou s parietálnymi chromatofórmi bez pyrenoidov.Medzi bunkami sú póry. Carpogon s trichogynom. Po oplodnení sa gonimoblasty vyvíjajú buď priamo z bruška oplodneného karpogónu, alebo z pomocných buniek po ich splynutí s oblasťovými vláknami. Nepohlavné rozmnožovanie je väčšinou pomocou tetraspór. Gonimoblasty nesúce karposporangiá sa považujú za špeciálnu generáciu - karposporofyt. Podrobnosti o vývoji karposporofytu (z oplodneného karpogónu alebo z pomocných buniek), ako aj čas diferenciácie pomocných buniek (pred alebo po oplodnení), ich poloha na stéli a pod. floridia do šiestich rádov 23
 Rad Nemalionaceae Zástupcovia sa vyznačujú absenciou pomocných buniek, gonimoblasty sa vyvíjajú priamo z oplodneného karpogónu 24
Rad Nemalionaceae Zástupcovia sa vyznačujú absenciou pomocných buniek, gonimoblasty sa vyvíjajú priamo z oplodneného karpogónu 24



 Rod Lemanea sa vyskytuje v rýchlo tečúcich riekach so studenou vodou. Talus má vzhľad nerozvetvených štetín 10-15 cm dlhých a 1 mm hrubých, tmavofialovej alebo olivovohnedej farby s nodulárnymi opuchmi. Pripevnené k podkladu pomocou podrážky vyrobenej z plazivých nití. Jedna niť predĺžených bezfarebných buniek prebieha pozdĺž osi talu. Z vrchu každej bunky stredovej osi sa rozprestiera klbko väčšinou štyroch lúčovitých vetiev. Bazálne bunky každej takejto vetvy sú veľké a predĺžené. Z ich distálneho (morfologicky horného) konca vybiehajú vetvy druhého rádu, ktoré sa zase rozvetvujú opakovane. Koncové vetvy zrastú do viacvrstvovej kôry. Jeho vonkajšie bunky sú malé a naplnené chromatofórmi, 28
Rod Lemanea sa vyskytuje v rýchlo tečúcich riekach so studenou vodou. Talus má vzhľad nerozvetvených štetín 10-15 cm dlhých a 1 mm hrubých, tmavofialovej alebo olivovohnedej farby s nodulárnymi opuchmi. Pripevnené k podkladu pomocou podrážky vyrobenej z plazivých nití. Jedna niť predĺžených bezfarebných buniek prebieha pozdĺž osi talu. Z vrchu každej bunky stredovej osi sa rozprestiera klbko väčšinou štyroch lúčovitých vetiev. Bazálne bunky každej takejto vetvy sú veľké a predĺžené. Z ich distálneho (morfologicky horného) konca vybiehajú vetvy druhého rádu, ktoré sa zase rozvetvujú opakovane. Koncové vetvy zrastú do viacvrstvovej kôry. Jeho vonkajšie bunky sú malé a naplnené chromatofórmi, 28
 Rad Cryptonemia obsahuje pomocné bunky, ktoré sa vyvíjajú pred oplodnením karpogónu a sú rozptýlené po celom talu v určitej vzdialenosti od karpogonu. Z karpogónu po oplodnení vyrastajú do pomocných buniek viac-menej dlhé mnohobunkové väzivové, čiže ooblastické vlákna. Po splynutí pomocnej bunky s bunkou ooblastického vlákna a prechode diploidného jadra vláknitej bunky do pomocnej bunky vznikajú gonimoblasty, ktoré sa vyvíjajú na gonimoblastoch (karposporofyty), karpospóry obsahujú diploidné jadro a vyklíčia na diploidné rastliny. - tetrasporofyty, ktoré produkujú len orgány nepohlavného rozmnožovania - tetrasporangia. Keď sa tvoria tetraspóry, dochádza k meióze a haploidné tetraspóry rastú do haploidných rastlín gametofytov nesúcich reprodukčné orgány. Gametofyt a tetrasporofyt sa navonok (morfologicky) nelíšia. izomorfná zmena generácií, komplikovaná diploidnou karposporoftózou. 29
Rad Cryptonemia obsahuje pomocné bunky, ktoré sa vyvíjajú pred oplodnením karpogónu a sú rozptýlené po celom talu v určitej vzdialenosti od karpogonu. Z karpogónu po oplodnení vyrastajú do pomocných buniek viac-menej dlhé mnohobunkové väzivové, čiže ooblastické vlákna. Po splynutí pomocnej bunky s bunkou ooblastického vlákna a prechode diploidného jadra vláknitej bunky do pomocnej bunky vznikajú gonimoblasty, ktoré sa vyvíjajú na gonimoblastoch (karposporofyty), karpospóry obsahujú diploidné jadro a vyklíčia na diploidné rastliny. - tetrasporofyty, ktoré produkujú len orgány nepohlavného rozmnožovania - tetrasporangia. Keď sa tvoria tetraspóry, dochádza k meióze a haploidné tetraspóry rastú do haploidných rastlín gametofytov nesúcich reprodukčné orgány. Gametofyt a tetrasporofyt sa navonok (morfologicky) nelíšia. izomorfná zmena generácií, komplikovaná diploidnou karposporoftózou. 29
 Durenea rod bežný v južných moriach. Talus vyzerá ako rozvetvené, slizké, ružové kríky. tridsať
Durenea rod bežný v južných moriach. Talus vyzerá ako rozvetvené, slizké, ružové kríky. tridsať
 31
31
 32
32
 33
33


 Rad Ceramiaceae je vysoko organizovaný, mnohé druhy pomocných buniek sa diferencujú až po oplodnení karpogónu Nevytvárajú sa dlhé vlákna: brucho karpogónu buď priamo splýva so susednou pomocnou bunkou (bunkami), alebo krátkymi procesmi , tvoriace cystokarpy 36
Rad Ceramiaceae je vysoko organizovaný, mnohé druhy pomocných buniek sa diferencujú až po oplodnení karpogónu Nevytvárajú sa dlhé vlákna: brucho karpogónu buď priamo splýva so susednou pomocnou bunkou (bunkami), alebo krátkymi procesmi , tvoriace cystokarpy 36


 evolúcia v rámci oddelenia Banguiaceae je primitívnejšia. Carpogon ešte nemá vyvinutú typickú formu a len málo sa líši od bežných vegetatívnych buniek. Po oplodnení sa obsah karpogónu priamo rozdelí na karpospóry. Floridai je viac vyvinutá skupina Carpogon s Trichogyna. Z triedy florídií je najjednoduchší rad nemalionaceae, ktoré nemajú pomocné bunky a priamo z brušnej časti oplodneného karpogónu vznikajú gonimoblasty, na ktorých sa vyvíjajú karposporangia. Kryptomiká sú ďalším krokom v progresívnej evolúcii: majú pomocné bunky, ktoré prispievajú k zvýšeniu produkcie karpospór, keďže sa netvorí jeden cystokarp, ale veľa - podľa počtu pomocných buniek. Keď sú pomocné bunky náhodne rozptýlené po celom taluse, sú potrebné viac alebo menej dlhé oblastic vlákna. Najvyšší stupeň evolúcie dosiahli ceramiidy, ktoré majú prokarp a pomocné bunky sa diferencujú až po oplodnení. Tesná blízkosť karpogónu a pomocnej bunky (buniek) v prokarpe uľahčuje tvorbu cpstokarpov. Tento rád je druhovo najbohatší. 39
evolúcia v rámci oddelenia Banguiaceae je primitívnejšia. Carpogon ešte nemá vyvinutú typickú formu a len málo sa líši od bežných vegetatívnych buniek. Po oplodnení sa obsah karpogónu priamo rozdelí na karpospóry. Floridai je viac vyvinutá skupina Carpogon s Trichogyna. Z triedy florídií je najjednoduchší rad nemalionaceae, ktoré nemajú pomocné bunky a priamo z brušnej časti oplodneného karpogónu vznikajú gonimoblasty, na ktorých sa vyvíjajú karposporangia. Kryptomiká sú ďalším krokom v progresívnej evolúcii: majú pomocné bunky, ktoré prispievajú k zvýšeniu produkcie karpospór, keďže sa netvorí jeden cystokarp, ale veľa - podľa počtu pomocných buniek. Keď sú pomocné bunky náhodne rozptýlené po celom taluse, sú potrebné viac alebo menej dlhé oblastic vlákna. Najvyšší stupeň evolúcie dosiahli ceramiidy, ktoré majú prokarp a pomocné bunky sa diferencujú až po oplodnení. Tesná blízkosť karpogónu a pomocnej bunky (buniek) v prokarpe uľahčuje tvorbu cpstokarpov. Tento rád je druhovo najbohatší. 39
 Najstaršia fosília identifikovaná ako červená riasa je zároveň najstaršou eukaryotickou fosíliou patriacou do špecifického moderného taxónu. Bangiomorpha pubescens, mnohobunková fosília nájdená v arktickej Kanade, sa len mierne líši od moderných červených rias rodu Bangia, napriek tomu, že bola uložená vo vrstvách pred 1,2 miliardami rokov. 41
Najstaršia fosília identifikovaná ako červená riasa je zároveň najstaršou eukaryotickou fosíliou patriacou do špecifického moderného taxónu. Bangiomorpha pubescens, mnohobunková fosília nájdená v arktickej Kanade, sa len mierne líši od moderných červených rias rodu Bangia, napriek tomu, že bola uložená vo vrstvách pred 1,2 miliardami rokov. 41
 DIVÍZIA HNEDÉ RIASY RNAEORNUTH morské, obzvlášť hojné v chladných vodách severnej a južnej pologule. Z hľadiska morfologickej a anatomickej diferenciácie talu je na vyššej úrovni ako doteraz uvažované skupiny rias. Nie sú známe ani jednobunkové ani koloniálne formy, ani stélka vo forme jednoduchého nerozvetveného vlákna. najjednoduchšie sú heterotrichné stélky, veľké, s falošnou alebo pravou tkanivovou štruktúrou.
DIVÍZIA HNEDÉ RIASY RNAEORNUTH morské, obzvlášť hojné v chladných vodách severnej a južnej pologule. Z hľadiska morfologickej a anatomickej diferenciácie talu je na vyššej úrovni ako doteraz uvažované skupiny rias. Nie sú známe ani jednobunkové ani koloniálne formy, ani stélka vo forme jednoduchého nerozvetveného vlákna. najjednoduchšie sú heterotrichné stélky, veľké, s falošnou alebo pravou tkanivovou štruktúrou.
 silne slizovité bunkové steny, jedno jadro, jedna alebo viac vakuol, zvyčajne stenové chromatofóry rôznych tvarov. chromatofóry sú obklopené zložitým systémom membrán - v priamom spojení s jadrovým obalom - „chloroplastové endoplazmatické retikulum“ Matricu chloroplastu pretínajú paralelné tritylakoidné lamely Pyrenoid bez tylakoidov vyčnieva z chloroplastu vo forme obličky
silne slizovité bunkové steny, jedno jadro, jedna alebo viac vakuol, zvyčajne stenové chromatofóry rôznych tvarov. chromatofóry sú obklopené zložitým systémom membrán - v priamom spojení s jadrovým obalom - „chloroplastové endoplazmatické retikulum“ Matricu chloroplastu pretínajú paralelné tritylakoidné lamely Pyrenoid bez tylakoidov vyčnieva z chloroplastu vo forme obličky
 Chromatoforové pigmenty sú sfarbené do hneda, chlorofyly „a“ a „c“ (chlorofyl „b“ chýba) β-karotén Existuje veľa hnedých xantofylov, najmä fukoxantín. 44
Chromatoforové pigmenty sú sfarbené do hneda, chlorofyly „a“ a „c“ (chlorofyl „b“ chýba) β-karotén Existuje veľa hnedých xantofylov, najmä fukoxantín. 44
 Mimo chloroplastu v cytoplazme sa ukladá rezervný polysacharid – laminarínový alkohol manitolové tuky. Monádové bunky (zoospóry a gaméty) majú oko a bičíky. Ocellus je súčasťou plastidu a je spojený s bičíkovým aparátom. Bičíky sú heteromorfné. 45
Mimo chloroplastu v cytoplazme sa ukladá rezervný polysacharid – laminarínový alkohol manitolové tuky. Monádové bunky (zoospóry a gaméty) majú oko a bičíky. Ocellus je súčasťou plastidu a je spojený s bičíkovým aparátom. Bičíky sú heteromorfné. 45


 Rozmnožovanie je vegetatívne, nepohlavné a sexuálne. Vegetatívne rozmnožovanie sekciami talu. Niektoré majú špecializované vetvy (plodové púčiky), ktoré sa lámu do nových stielok. Nepohlavné rozmnožovanie - zoospóry vznikajú v jednobunkových alebo jednokomorových výtrusniciach na diploidných rastlinách (sporofytoch) pred tvorbou zoospór, jadro je redukovane rozdelené Haploidné zoospóry - na haploidné pohlavné rastliny - gametofyty, na ktorých sa tvoria pohlavné orgány. U prvokov je pohlavný proces izogamný, gaméty sa vyvíjajú v multilokulárnych alebo viackomorových sporangiách.U najorganizovanejších hnedých rias je pohlavný proces oogamný. V oogónii a anterídii sa spravidla tvorí jedna gaméta (vajíčko a spermie). Vajíčko je vždy oplodnené mimo oogium. Zygota rastie do diploidnej rastliny bez obdobia vegetačného pokoja.
Rozmnožovanie je vegetatívne, nepohlavné a sexuálne. Vegetatívne rozmnožovanie sekciami talu. Niektoré majú špecializované vetvy (plodové púčiky), ktoré sa lámu do nových stielok. Nepohlavné rozmnožovanie - zoospóry vznikajú v jednobunkových alebo jednokomorových výtrusniciach na diploidných rastlinách (sporofytoch) pred tvorbou zoospór, jadro je redukovane rozdelené Haploidné zoospóry - na haploidné pohlavné rastliny - gametofyty, na ktorých sa tvoria pohlavné orgány. U prvokov je pohlavný proces izogamný, gaméty sa vyvíjajú v multilokulárnych alebo viackomorových sporangiách.U najorganizovanejších hnedých rias je pohlavný proces oogamný. V oogónii a anterídii sa spravidla tvorí jedna gaméta (vajíčko a spermie). Vajíčko je vždy oplodnené mimo oogium. Zygota rastie do diploidnej rastliny bez obdobia vegetačného pokoja.

 TRIEDA ISOGENERATES Objednávka Ectocarpous Rod Ectocarpus - najprimitívnejšia hnedá riasa. Sú rozšírené vo všetkých moriach, najmä studených, rastú na podvodných predmetoch a iných väčších riasach. 50
TRIEDA ISOGENERATES Objednávka Ectocarpous Rod Ectocarpus - najprimitívnejšia hnedá riasa. Sú rozšírené vo všetkých moriach, najmä studených, rastú na podvodných predmetoch a iných väčších riasach. 50
 51
51
 52
52

 54
54
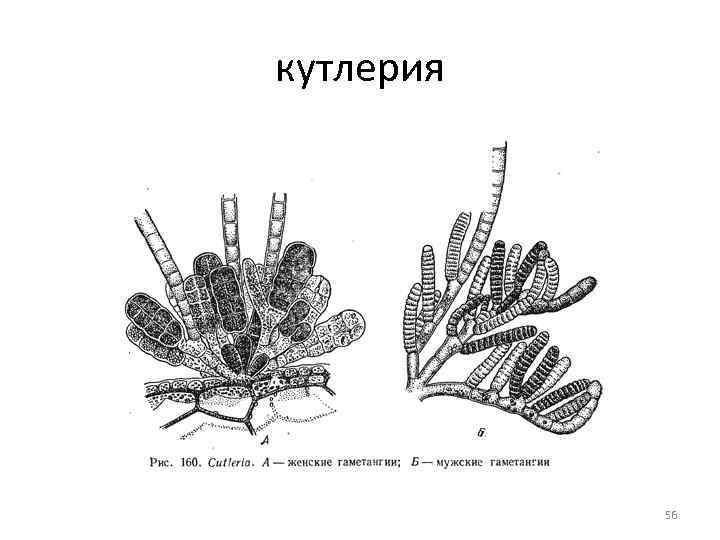
 Rad Cutleriaceae Rad zahŕňa len dva rody: Cutleria a Zanardinia Cutleria je rozšírená pozdĺž celého pobrežia Európy, Zanardínie - hlavne v Stredozemnom mori, ako aj v Čiernom mori. Oba rody vykazujú striedanie generácií: v Cutleria je zmena generácií heteromorfná, v Zanardínii je to izomorfná 55
Rad Cutleriaceae Rad zahŕňa len dva rody: Cutleria a Zanardinia Cutleria je rozšírená pozdĺž celého pobrežia Európy, Zanardínie - hlavne v Stredozemnom mori, ako aj v Čiernom mori. Oba rody vykazujú striedanie generácií: v Cutleria je zmena generácií heteromorfná, v Zanardínii je to izomorfná 55
 57
57
 58
58

 60
60

 62
62
 TRIEDA HETEROGENERÁT Rad Laminaria Gametofyty sa navzájom málo líšia a sú zastúpené mikroskopickými, často na niekoľko buniek redukovanými, vláknitými rastlinkami, ktoré nesú rozmnožovacie orgány. Na samčích gametofytoch sa tvoria anterídie vo forme malých buniek, ktoré vyvíjajú jednu spermiu, na samičích gametofytoch - oogóniách, v ktorých sa tvorí jedno vajíčko. Sporofyty rôznych rodov sa výrazne líšia a predstavujú najväčšie nižšie rastliny, dosahujúce dĺžku 60-100 m, s výrazným morfologickým členením a zložitou anatomickou stavbou. Vo väčšine prípadov sú sporofyty riasy rozdelené na čepeľ v tvare listu, „kmeň“ a rizoidy, pomocou ktorých je celá rastlina pripevnená k podvodným kameňom a skalám. Na styku čepele listu a stonky sa nachádza interkalárny meristém, vďaka činnosti ktorého čepeľ listu aj stonka rastú. rod chaluhy, ktorej druhy sú rozšírené v severných moriach. 63
TRIEDA HETEROGENERÁT Rad Laminaria Gametofyty sa navzájom málo líšia a sú zastúpené mikroskopickými, často na niekoľko buniek redukovanými, vláknitými rastlinkami, ktoré nesú rozmnožovacie orgány. Na samčích gametofytoch sa tvoria anterídie vo forme malých buniek, ktoré vyvíjajú jednu spermiu, na samičích gametofytoch - oogóniách, v ktorých sa tvorí jedno vajíčko. Sporofyty rôznych rodov sa výrazne líšia a predstavujú najväčšie nižšie rastliny, dosahujúce dĺžku 60-100 m, s výrazným morfologickým členením a zložitou anatomickou stavbou. Vo väčšine prípadov sú sporofyty riasy rozdelené na čepeľ v tvare listu, „kmeň“ a rizoidy, pomocou ktorých je celá rastlina pripevnená k podvodným kameňom a skalám. Na styku čepele listu a stonky sa nachádza interkalárny meristém, vďaka činnosti ktorého čepeľ listu aj stonka rastú. rod chaluhy, ktorej druhy sú rozšírené v severných moriach. 63
 TRIEDA CYCLOSPORONAE Rad Fucus sa vyznačuje - - apikálnym rastom talu.Neprítomnosť nepohlavného rozmnožovania oogamným pohlavným procesom.Pohlavné orgány sa nachádzajú v priehlbinách talu - pojmoch,alebo scafidia.Existuje vegetatívne rozmnožovanie, vedúce v niektorých častiach oceánu k vzniku obrovských akumulácií Sargassum thalli (príkladom je Sargasové more).
TRIEDA CYCLOSPORONAE Rad Fucus sa vyznačuje - - apikálnym rastom talu.Neprítomnosť nepohlavného rozmnožovania oogamným pohlavným procesom.Pohlavné orgány sa nachádzajú v priehlbinách talu - pojmoch,alebo scafidia.Existuje vegetatívne rozmnožovanie, vedúce v niektorých častiach oceánu k vzniku obrovských akumulácií Sargassum thalli (príkladom je Sargasové more).
 rod Sargassum komplexné morfologické členenie talu je rozšírené na južnej pologuli, najmä v teplých moriach. Stonka je prichytená k substrátu podošvou, konáre, nesie sploštené listovité útvary, guľovité vzduchové bubliny na zvláštnych stopkách, rozvetvené plodné konáre 71
rod Sargassum komplexné morfologické členenie talu je rozšírené na južnej pologuli, najmä v teplých moriach. Stonka je prichytená k substrátu podošvou, konáre, nesie sploštené listovité útvary, guľovité vzduchové bubliny na zvláštnych stopkách, rozvetvené plodné konáre 71



 Hnedé riasy sú prirodzenou skupinou, ktorá sa svojou morfológiou dobre odlišuje od ostatných oddelení rias. Ako však už bolo spomenuté, podľa viacerých charakteristík (prítomnosť chlorofylov „a“ a „c“, neprítomnosť chlorofylu „b“, umiestnenie tylakoidov v chloroplastoch – tritylakoidné lamely, podľa podobných skladovacích produktov – chaluha , chrysolamín, podľa štruktúry monadických buniek s heterokontom a heteromorfným bičíkom) hnedé riasy vykazujú podobnosti so zlatými, žltozelenými, rozsievkami a pyrofytickými riasami. Na tomto základe niektorí autori zaraďujú hnedé a iné uvedené skupiny rias do triedy tried v rámci jednej veľkej divízie Credoshore. bу1 a. Existencia monadických buniek v životnom cykle hnedých rias umožňuje, aby boli rovnako ako ostatné uvedené skupiny odvodené od niektorých primárnych fotosyntetických bičíkovcov s prevahou hnedých pigmentov. Tieto bičíkovce sa vyvinuli niekoľkými spôsobmi, z ktorých jeden viedol k vzniku hnedých rias. Hnedé riasy však majú s bičíkovcami vzdialenejšie spojenie ako zlaté riasy, heterobičíkovce, rozsievky, pyrofyty a zelené riasy, ktoré sú si v mnohých vlastnostiach podobné, ako aj zelené riasy, keďže tu neexistujú žiadne priame prechodné formy. Medzi hnedými riasami neexistuje konzistentný vývoj typov organizácie talu od monadických po vláknité a lamelárne, ako je ľahko pozorovateľné u zelených, žltozelených, zlatých a pyrofytických rias. Ako už bolo uvedené, medzi hnedými riasami sú zastúpené iba najvyššie štádiá morfologickej diferenciácie talu - heterotrichálne a lamelárne. Možno, že jednoduchšie formy (monáda, kokoid, jednoduché vláknité) sa úplne stratili počas dlhého vývoja hnedých rias - starodávnej už známej skupiny; zo silúrskych a devónskych ložísk. Neexistuje jednotný pohľad na súvisiace vzťahy objednávok v rámci oddelenia hnedých rias a ich umiestnenie v systéme. Podľa jednej zo schém fylogenézy hnedých rias, skonštruovaných s prihliadnutím na podobnosť štruktúry stéla v najskorších štádiách vývoja, hnedé riasy sú difyletickou skupinou: jedna línia evolúcie sa spája (z tých, ktoré sú uvedené v prezentácii ) Sphacelariaceae, Dictyotaceae a Fucus, ostatné - ektocarpaceae, Cutleriaceae a Laminariaceae. Oba 219" 75
Hnedé riasy sú prirodzenou skupinou, ktorá sa svojou morfológiou dobre odlišuje od ostatných oddelení rias. Ako však už bolo spomenuté, podľa viacerých charakteristík (prítomnosť chlorofylov „a“ a „c“, neprítomnosť chlorofylu „b“, umiestnenie tylakoidov v chloroplastoch – tritylakoidné lamely, podľa podobných skladovacích produktov – chaluha , chrysolamín, podľa štruktúry monadických buniek s heterokontom a heteromorfným bičíkom) hnedé riasy vykazujú podobnosti so zlatými, žltozelenými, rozsievkami a pyrofytickými riasami. Na tomto základe niektorí autori zaraďujú hnedé a iné uvedené skupiny rias do triedy tried v rámci jednej veľkej divízie Credoshore. bу1 a. Existencia monadických buniek v životnom cykle hnedých rias umožňuje, aby boli rovnako ako ostatné uvedené skupiny odvodené od niektorých primárnych fotosyntetických bičíkovcov s prevahou hnedých pigmentov. Tieto bičíkovce sa vyvinuli niekoľkými spôsobmi, z ktorých jeden viedol k vzniku hnedých rias. Hnedé riasy však majú s bičíkovcami vzdialenejšie spojenie ako zlaté riasy, heterobičíkovce, rozsievky, pyrofyty a zelené riasy, ktoré sú si v mnohých vlastnostiach podobné, ako aj zelené riasy, keďže tu neexistujú žiadne priame prechodné formy. Medzi hnedými riasami neexistuje konzistentný vývoj typov organizácie talu od monadických po vláknité a lamelárne, ako je ľahko pozorovateľné u zelených, žltozelených, zlatých a pyrofytických rias. Ako už bolo uvedené, medzi hnedými riasami sú zastúpené iba najvyššie štádiá morfologickej diferenciácie talu - heterotrichálne a lamelárne. Možno, že jednoduchšie formy (monáda, kokoid, jednoduché vláknité) sa úplne stratili počas dlhého vývoja hnedých rias - starodávnej už známej skupiny; zo silúrskych a devónskych ložísk. Neexistuje jednotný pohľad na súvisiace vzťahy objednávok v rámci oddelenia hnedých rias a ich umiestnenie v systéme. Podľa jednej zo schém fylogenézy hnedých rias, skonštruovaných s prihliadnutím na podobnosť štruktúry stéla v najskorších štádiách vývoja, hnedé riasy sú difyletickou skupinou: jedna línia evolúcie sa spája (z tých, ktoré sú uvedené v prezentácii ) Sphacelariaceae, Dictyotaceae a Fucus, ostatné - ektocarpaceae, Cutleriaceae a Laminariaceae. Oba 219" 75
 Skupiny rádov sa líšia aj spôsobom rastu stél: prvý sa vyznačuje apikálnym rastom, druhý interkalárnym rastom. Kilin (N. Ku 1sh), ktorého systém je v súčasnosti najrozšírenejší, odvodzuje od spoločného predka hnedých rias tri evolučné línie, ktoré prezentuje ako izogenerátne, heterogenerátne a cyklosporózne triedy; sú založené na rozdieloch v životných cykloch a morfologickej štruktúre rias. Veľké hnedé riasy sú široko používané v ľudskom poľnohospodárstve. Extrahuje sa z nich algin – lepiaca látka používaná v textilnom, potravinárskom a množstve ďalších odvetví. V pobrežných krajinách sa emisie morských rias, bohaté na draslík a dusík, používajú ako hnojivo a tiež ako krmivo pre dobytok. Niektoré, ako napríklad kelp (morské riasy), sú jedlé. 76
Skupiny rádov sa líšia aj spôsobom rastu stél: prvý sa vyznačuje apikálnym rastom, druhý interkalárnym rastom. Kilin (N. Ku 1sh), ktorého systém je v súčasnosti najrozšírenejší, odvodzuje od spoločného predka hnedých rias tri evolučné línie, ktoré prezentuje ako izogenerátne, heterogenerátne a cyklosporózne triedy; sú založené na rozdieloch v životných cykloch a morfologickej štruktúre rias. Veľké hnedé riasy sú široko používané v ľudskom poľnohospodárstve. Extrahuje sa z nich algin – lepiaca látka používaná v textilnom, potravinárskom a množstve ďalších odvetví. V pobrežných krajinách sa emisie morských rias, bohaté na draslík a dusík, používajú ako hnojivo a tiež ako krmivo pre dobytok. Niektoré, ako napríklad kelp (morské riasy), sú jedlé. 76
- "úžasný" kongres národov Tatarstanu
- Medzietnický konflikt v Dagestane sa skončil víťazstvom Kadyrova: názor Kto teraz žije v tejto oblasti
- Archív konferencií a seminárov
- Budete hovoriť o súhlase rodičov, aby ste sa naučili svoj rodný jazyk
- Rustem Khamitov oznámil možné zrušenie baškirského jazyka v školách republiky Vyučuje sa baškirský jazyk v Baškirsku?
- Didaktický materiál pre GIA v ruskom jazyku Vykonávanie testu samostatná práca
- Účel: študovať výsledky ciest prezentácie Jamesa Cooka na tému James Cook